ERCIM News 64January 2006Contents 
This issue in pdf Subscription Archive: Next issue: April 2006 Next Special theme:
|
by Heather Ruskin, and John Burns
Individual experiences of disease can be extremely diverse. As a result, if targeted treatment of the individual is to be achieved, we need some way of assessing the variables that lead to different outcomes for different people. Visualizing the record of viral attacks in terms of a network of connections that are formed and broken in defence against an attack, can aid in understanding observed effects. This is similar to mapping and redrawing operational outcomes on a larger scale, in order to incorporate necessary or unforeseen changes. Here we describe some of the ideas involved in the formation of connected networks of information at the cellular level, where these provide immune system fingerprints of individual exposure.
Emergent principles of the immune system T-cell repertoire and its self-organization can be simply modelled for a lymphatic compartment, using stochastic cellular automata. The immune system hybrid model incorporates a shape space formalism, permitting each activated effector T-cell clonotype and viral epitope to be represented as nodes of a graph, with edges modelling the affinity or clearance pressure applied to the antigen-presenting cell (APC,) which bears the target epitope. For repeated exposure to infection by heterologous or mutating viruses, a distinct topology of the network space emerges. Properties of the emergent network reflect recent experimental results on cytotoxic T-cell (CTL) activation, apoptosis, cross-reactivity and memory, especially with respect to re-infection.
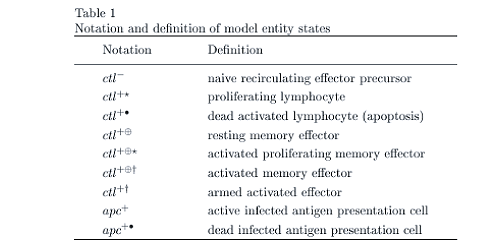
Biological systems are complex by nature, and the immune system is typical in this respect. In the hybrid model outlined, the real space of the lymphatic compartment - with entities and states (cell-types) as in Table 1 - exchanges information with the shape space, which models affinity between cell types and clearance pressure. The CTL population grows exponentially in response to APC stimulation, with a rate dependent on the distance between the APC and CTL in shape space. The model permits:
Emergent behaviour is thus explicitly exhibited.
|
The focus here is on two cell- types only, APC and CTL. We omit discussion of the humoral response as well as reference to CD4+ T-cells - their role in facilitating CTL activity is implicitly assumed. Dynamics, governing the affinity between antigen and lymphocyte, arise from the shape space formalism (see Perelson A.S. and Oster G.F. (1979), Theoretical Studies of Clonal Selection: Minimal Antibody Repertoire Size and Reliability of Self-Non-Self Discrimination. J. Theor. Biol., 81 (4):645-70). Each unique antigen epitope and CTL clone is represented as a point within the two-dimensional space of given size. Surrounding each CTL clone is a disc of radius r. Any antigen epitope located within the disc is subjected to a clearance pressure with a force inversely proportional to the separation distance (see Burns J. and Ruskin H.J. (2004) Diversity Emergence and Dynamics during Primary Immune Response: A Shape Space, Physical Space Model: Theor. in Biosci., 123(2):183-194). A network model of shape space emerges naturally from the real-space model, with each immunogenic epitope ek and activated CTL clonotype cj represented by a node in the space. Clearance pressure applied between two nodes forms a directed edge between them, with a weight dependent on the affinity. After initial infection, most cj undergo programmed apoptosis (a crucial regulator of immune system homeostasis), which corresponds to edge deletion. However, recruitment to the memory pool consumes a given percentage of activated CTL, so nodes remain active in shape space, preserving edge connections to stimulatory epitopes.
Recent work indicates that effector CTL memory cells can recognise epitopes of unrelated viruses, so that heterologous viruses (derived from a separate genetic source) may be a key factor in influencing the hierarchy of CD8+ T-cell responses and the shape of memory T-cell pools. Shape space can be used to model both homogeneous viruses with conserved and mutated epitopes, and homologous viruses, with cross-reactive epitopes.
|
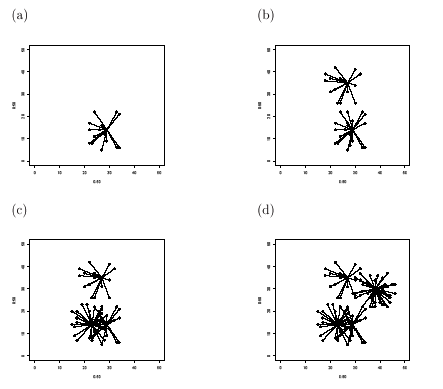
Early and protective immunity can be mediated by memory T-cells generated by previous heterologous infection (represented graphically by nodes with degree > 2). Cluster linkage illustrates conditions by which immunity to one virus can reduce the effects of challenge by another (see Figure 1). Damage to or suppression of critical cross-reactive 'a'-nodes has significantly greater impact than damage to leaf or 'b'-nodes. Different disease outcomes to identical infection strains can be explained in terms of density and distribution of a-nodes. The degree of protection from cross-reacting memory cells depends on the distance between the memory T-cell clonotype and immunogenic epitope, with optimal immunity for re-infection by the same antigenic epitope. Results show increasingly effective clearance dynamics as the memory pool increases and each T-cell clone has a finite chance of becoming a long-lived memory cell.
Please contact:
Heather J. Ruskin, Dublin City University / IUA, Ireland
E-mail: hruskin![]() computing.dcu.ie
computing.dcu.ie